
Unser Körper ist aus zirka 13 Billionen Zellen aufgebaut. Jede Zelle (Abb.3) stellt einen kleinen Organismus dar, der in der Lage ist, mit seinen benachbarten Zellen und/oder der Außenwelt des Gesamtorganismus zu kommunizieren. Im Gegensatz zu den Einzellern, die alle biologischen Funktionen gleichzeitig bewerkstelligen müssen, finden wir bei den Vielzellern eine mehr oder weniger ausgeprägte funktionelle Differenzierung und Spezialisierung der einzelnen Zelltypen. Man zählt heute mehr als 250 funktionell verschiedene Zellarten. Wir werden später sehen, daß alle Zellen eines Individuums die gleichen genetischen Informationen tragen, die jedoch entsprechend ihrem Differenzierungsgrad oder ihrem Funktionszustand einzeln oder gruppenweise „an- oder abgeschaltet“ werden können.
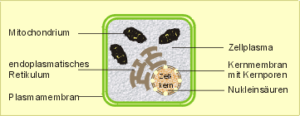
Abb. 3: Schematischer Aufbau einer Zelle
Zellmembran. Beginnen wir mit der Zellmembran oder Plasmamembran, die den Zellinnenraum umschließt und die Schranke zum extrazellulären Raum bildet. Hier finden alle stofflichen und Informationsaustausche mit der „inneren Umwelt“ statt, die der Erhaltung, dem Wachstum und der Reproduktion der Zelle dienen. Dementsprechend ist ihr Bau sehr differenziert und hochdynamisch. Die Zellmembran bildet in organisierten Geweben die Grenze zu den Nachbarzellen und im Fall „flüssiger Gewebe“ wie etwa dem Blut oder der Lymphe die Schranke zum umgebenden Flüssigkeitsstrom.
Die elekronenmikroskopisch sichtbare Grundstruktur der Plasmamembran ist eine Doppelmembran, die für sich allein genommen, ziemlich inert ist und kaum zur Ausführung von fein abgestimmten Regelprozessen geeignet wäre. Sie besteht zur Hälfte aus verschiedenen Lipidmolekülen (z.B. Cholesterin, Phospholipide) und zur anderen Hälfte aus Proteinen oder Abkömmlingen von reinen Proteinen (z.B. Glykoproteinen, Lipoproteinen). Der Fachterminus Zellwand wird im Gegensatz zur Plasmamembran für jene Schichten verwendet, die bei der Bakterien- und Pflanzenzelle der Plasmamembran zusätzlich aufgelagert sind und hauptsächlich aus Zuckermolekülen und deren komplexen Abkömmlingen bestehen (Abb. 4).


Abb. 4: Die tierische Zelle wird durch eine Doppelmembran begrenzt. Der Pflanzenzelle ist zusätzlich eine kohlenhydratreiche Zellwand aufgelagert
An der Oberfläche der Plasmamembran läßt sich eine Vielzahl von verschiedenen Makromolekülen – meist Glykoproteine – nachweisen, deren Funktion bisher nur unvollständig aufgeklärt ist. Sie bilden in Abhängigkeit von der funktionellen Differenzierung (von der Stammzelle zur Organzelle) und dem aktuellen funktionellen Zustand charakteristische Molekülmuster (vgl. Abb. 37; Abschnitt „Antigene – Antikörper“). Einige dieser Zelloberflächenmoleküle jedoch sind individualspezifisch, d.h. ihre Struktur unterscheidet sich von Individuum zu Individuum. Sie bilden z.B. die Grundlage für die Blutgruppen- und Gewebetypisierung und die Risikoabschätzung für verschiedene Krankheiten wie den verschiedenen Leukämien. Die erwähnten Regelprozesse an der Zellmembran werden von Makromolekülen – meist Eiweißen – bewerkstelligt, die der Membran innen oder außen auf- oder eingelagert sind oder als Transmenbranmoleküle den extrazellulären mit dem intrazellulären Raum verbinden. Ohne uns weiter in diese hochinteressante Materie zu vertiefen, soll noch erwähnt werden, daß die Membranproteine nicht in der Membran starr fixiert sind, sondern daß die Plasmamembran aufgrund ihrer Struktur als flüssiger Kristall (denken Sie an die LCD-Anzeige in ihrem Taschenrechner) die Bewegung der Membranbranproteine in der Membran zulassen – sie „schwimmen“ in der Membran.
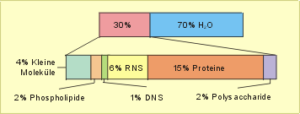
Zytoplasma. Kommen wir zum Zellinnenraum. Er wird ausgefüllt vom Zytoplasma, einem Gemisch von Salzen und Makromolekülen (Abb. 5) sowie von geformten, mikroskopisch oder elektronenmikroskopisch darstellbaren Zellorganellen. Wichtig zu wissen ist, daß der Zellinnenraum keine amorphe Suppe darstellt, sondern wohlorganisiert ist und „alles seinen Platz hat“. Dafür sorgen Membran- und Faserstrukturen, die die einzelnen funktionellen Kompartimente voneinander abtrennen und die Beziehungen zwischen ihnen organisieren.
Im folgenden sollen nun einige Zellorganelle besprochen werden, soweit sie zum Verständnis der weiteren spezielleren Ausführungen erforderlich sind. Dies sind: der Zellkern, das endoplasmatische Retikulum mit den Ribosomen und die Mitochondrien.
Zellkern. Beim Menschen haben – bis auf die reifen roten Blutkörperchen – alle Zellen einen Zellkern. Der Kern beherbergt den größten Teil der genetischen Informationen, die über die Keimzellen an die Nachfahren weitergegeben werden. Sie ist in den Nukleinsäuren (DNS, RNS) gespeichert und in den Chromosomen lokalisiert. Nur während einer bestimmten Phase der Zellteilung (Metaphase) sind die Chromosomen nach Anfärbung sichtbar.
Auch der Zellkern wird wie die gesamte Zelle von einer Doppelmembran umgeben. Er besitzt Kernporen, die während der Eiweiß-Biosynthese eine wichtige Rolle beim Transport und der Übersetzung der im Kern gespeicherten genetischen Information in die Aufeinanderfolge der Eiweißbausteine (Aminosäuren) spielt (vgl. Abschnitt „Vom Gen zum Eiweiß“). Die Kernporen sind für die riesigen DNS-Moleküle undurchlässig, jedoch für die kleineren Überträger der genetischen Information, die Messenger-RNS (Boten-RNS), durchlässig. Auf diese Weise können letztere in den Zellinnenraum gelangen, wo sich in der Nähe des Zellkerns die Biosynthese der Eiweiße abspielt. Dieses endoplasmatische Retikulum besteht aus einem Netzwerk von Doppelmembranen (endoplasmatisches Retikulum), denen die sog. Ribosomen aufgelagert sind (daher der Name „rauhes“ endoplasmatisches Retikulum). Hier findet die Eiweißsynthese, also der Übergang der genetischen Informationen in die Primärstruktur der Eiweiße, statt.
Mitochondrien. Die Mitochondrien sind stäbchenförmige bis kugelige Organelle, die als „Kraftwerke der Zelle“ bezeichnet werden, da sie als der Hauptort der Biosynthese von universellen Energieträgern (ATP) angesehen werden, die an den meisten energieabhängigen Stoffwechselreaktionen beteiligt sind. Im Zusammenhang mit unserem Thema ist zu erwähnen, dass die Matrix der Mitochondrien ringförmige DNS und RNS sowie Ribosomen enthält, also den gesamten prinzipiellen Informations- und Syntheseapparat, wie er für die chromosomale DNS und RNS sowie das endoplasmatische Retikulum für den Zellkern soeben beschrieben wurde. So ist es nicht überraschend, dass es neben der chromosomalen Vererbung auch eine mitochondriale Vererbung gibt, die allerdings auf eine Weitergabe der Erbinformationen in weiblicher Linie beschränkt ist (vgl. Abschnitt „Das Mitochondriengenom“).
Fazit: Die genetische Information ist im Zellkern und den Mitochondrien gespeichert. Die Übersetzung der genetischen Information in Eiweißstrukturen findet außerhalb des Zellkerns in der Nähe der Kernmembran statt.