
Kehren wir noch für einen Moment zum Genom zurück. Wir hatten festgestellt, daß es sich beim Genom trotz der hauptsächlich durch Mutation und Rekombination bedingten relativen Variabilität um eine recht stabile Struktur handelt, die in mehr oder weniger unveränderter Form an die Nachkommen weitergegeben wird. Wie aber verhält sich das Genom zu den Produkten (Eiweiße), die nach den Informationen des genetischen Bauplans synthetisiert werden? Es geht also um die Frage nach dem Verhältnis zwischen Genotyp und Phänotyp. Eines kann schon vorweg gesagt werden: Aus den einzelnen Nukleotidsequenzen kann nicht unmittelbar auf die Endprodukte geschlossen werden. Viele Gene sind nämlich fragmentiert; andere, sogenannte Pseudogene, sind unvollständig, und eine nicht unbeträchtliche Zahl liegt mitunter weit über das Genom verteilt in mehreren Kopien vor.
Die primäre Regulation der Aktivität der meisten Gene findet auf der Stufe der Transkription statt. Bereits bei der ersten Ablesung der DNS und ihrer Kopierung in eine Boten-RNS gibt es regulative Einflüsse, die dem endgültigen Eiweißmolekül veränderte strukturelle und folglich auch funktionelle Eigenschaften verleihen können. Wir hatten erfahren, daß der erste Schritt der Transkription in der Herstellung einer instabilen Kopie der DNS in Form einer prä-mRNS besteht, aus der in einem nächsten Schritt durch das sog. Spleißen die nichtkodierenden Introns enzymatisch herausgeschnitten werden. Beim „Zusammensetzen“ der kodierenden Exons zur endgültigen Boten-RNS kann es insbesondere beim sogenannten alternativen Spleißen zu Nukleotidsequenzen kommen, die etwas von der ursprünglichen Sequenz abweichen. Dies führt, wie zu erwarten, auch zu abweichenden Proteinen und äußert sich meist in einer unterschiedlichen räumlichen Proteinstruktur und zwangläufig in neuen Wechselwirkungsmöglichkeiten dieser Moleküle. Dies muß für den Organismus nicht von Schaden sein, sondern ist z.B. bei der Bildung spezifischer Antikörper im Verlauf der Immunreaktion offensichtlich eine notwendige Voraussetzung (vgl. Abschnitt „Antigene-Antikörper„).
Den größten Einfluß darauf, welche Gene, wann und wie lange abgelesen werden, also „aktiv“ sind, wird vordergründig vom lokalen Milieu, das das genetische Material umgibt, bestimmt. Milieu bedeutet in diesem Zusammenhang die Zusammensetzung des Kernplasmas und der anderen „Kompartimente“ der Zelle sowie die Konzentration und Konformation der einzelnen Moleküle, die auf den Prozess der einzelnen Stufen der Eiweiweißsynthese regulativ einwirken. So herrscht in einer Leberzelle ein anderes Milieu als in einer Nervenzelle und die Art und Zahl der produzierten Eiweiße wird sich wesentlich unterscheiden. Diese Systeme sind hochdynamisch und meist in große Regelkreise und biochemische Netzwerke eingebunden. Zellteilung, Wachstum, Biorhythmus, selbst der programmierte Zelltod unterliegen einer zweckentsprechenden Regulation der Genaktivität.
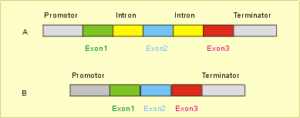
Sehen wir uns ein Gen und seine Umgebung etwas genauer an (Abb. 14). Im Abschnitt „Vom Gen zum Eiweiß“ wurde bereits gezeigt, daß ein Gen prinzipiell aus kodierenden Exons und nichtkodierenden Introns besteht. Beginn und Ende der kodierenden Abschnitte werden durch jeweils ein für die Ablesung wichtiges Start- und ein Stopkodon begrenzt. Unmittelbar vor der Startregion befindet sich die sogenannte Promotorregion, eine der wichtigen Ansatzstellen für regulative Moleküle (z.B. Hormone), die für das An- und Abschalten der Ablesung der genetischen Information (Transkription) verantwortlich sind. Manchmal weit vom Transkriptionsstartpunkt entfernt, können außerdem kurze Basensequenzen (sog. Enhancer [Verstärker] bzw. Silencer [Abschwächer]) liegen, die die Ablesung eines DNS-Abschnitts herauf- oder herunterregeln. Sie bilden die Andockstelle für verschiedene Regulatorproteine. Enhancer und Silencer wirken in bestimmten Geweben, zu bestimmten Zeiten oder in Anwesenheit bestimmter Signale.
Von den oben beschriebenen Genen können sogenannte Housekeeping-Gene unterschieden werden. Das sind Gene, die nicht reguliert und unabhängig von Umwelteinflüssen in allen Zellen konstant exprimiert werden. In der Regel kodieren sie Enzyme, die der Aufrechterhaltung des Grundtoffwechsels dienen sowie Proteine des Zytoskeletts, ribosomale RNA und Histone.
Fazit: Die Übersetzung der genetischen Information in Eiweißstrukturen wird durch mannigfaltige Regulationsmechanismen beeinflußt. Was, wann, wie, wo und wie lange „angeschaltet“ und übersetzt wird, hängt im dynamischen Geschehen vom molekularen Umfeld (Milieu) ab, das das genetische Material umgibt. Molekulare Ansatzpunkte (Promotoren) befinden sich vor den Startbereichen der Ablesung eines Gens bzw. weiter entfernt in Form von Nukleotidsequenzen zum Herauf- und Herrunterregeln (Enhancer und Silencer).